Sidebar. Decline of nutrient content in plants at high CO2, particularly nitrogen
“When I was a boy,” as the old line goes… the quoted molar mixing ratio of CO2 in the atmosphere was 300 parts per million (ppm). As of end June 2020, it is at least 414 ppm, a gain of 38% with no sign of slowing down except a bit from the COVID-19 pandemic reducing transportation use of fossil fuels. Now, CO2 and water are the main substrates for plant growth: CO2* + 2 H2O [CH2O] + H2O*], where the asterisk indicates that oxygen in the new water is from CO2, and the bracketed entity is 1/6 of a glucose molecule.
Photosynthesis as limited by the rubisco enzyme. All else equal, the photosynthetic rate of a leaf of a given composition should rise close to proportionally to the level of CO2. The rate-limiting step in photosynthesis is the carboxylation of ribulose 1,5-bisphosphate, catalyzed by the enzyme known by its short name, rubisco (also a good name for a dog, I think; “Here, rubisco, come, rubisco”) or in full as ribulose bisphosphate carboxylase oxygenase. The rate of carboxylation vs. CO2 partial pressure inside the leaf has the form below at saturatingly high light intensity (typical result, in this case from VanLeeuwen et al., 2014; pecan tree):
Leaves respond to many factors (cf. Gutschick, 2007), particularly CO2, humidity, temperature, water status, and light in the short term. Carbon dioxide enters the leaf interior and water vapor exits through exquisitely controlled pores called stomata. If CO2 varies, the common result of stomatal control for a plant in sufficient light intensity (photosynthetic photon flux density and not under water stress is to maintain internal CO2 levels at a nearly constant fraction of CO2 in outside air, about 70%. A plant at 300 ppm CO2 has an internal level of about 210 ppm, and one at 414 ppm has an internal level of about 283 ppm (all this should really be in pascals, but that’s not critical for this argument). From the curve above we may deduce an expected increase in photosynthetic rate from 2.25 to 3.01 in the units noted as CO2 increases from the lower level to the higher level. That’s an increase of 34%, just about the same proportion as in CO2 itself.
However, plants adapt to high CO2. This has been the subject of an enormous amount of scientific research for agronomy, ecology, physiology, and climate science. A good part of my own research has been on this response and on the response to varying availability of nutrients. Plants subjected to high CO2 undergo significant changes in growth rate, photosynthetic capacity, hydraulic properties, allocation of growth to roots vs. shoots, and more. They generally lower their nutrient content, especially of nitrogen. This is readily viewed as adaptive. The main use of nitrogen is in leaves, and the main use in leaves is for photosynthetic enzymes (and chlorophyll). The main investment in enzymes is in rubisco because it has low rates of action; thus, it’s needed in much higher amounts than any other enzyme in the photosynthetic system. The low activity is a consequence of the biochemical difficulty of the reaction, second only to the reaction of nitrogen fixation. In any event, as CO2 inside the leaf increases, the efficacy of nitrogen in rubisco increases. For the same rate of photosynthesis per unit leaf area, a leaf could use notably less nitrogen in rubisco. For the case cited, the N content could be reduced by a factor 1/1.34 or 75%.
Plants don’t maintain the same rate of photosynthesis. They do have an increased rate but they reduce their N content partially, not in the proportion above. The gain in efficacy of nitrogen is tempered in the evolutionary term by the general difficulty of acquiring nitrogen from soil; it’s often the most limiting nutrient. The argument developed by Lou Ellen Kay and me is one of functional balance between roots (getting N) and leaves (using N in photosynthesis). All else equal (is that ever really true?), the content of N in plants varies roughly as the square root of the efficacy of N. Here, efficacy is measured as photosynthetic rate per mass of nitrogen.
We explored the functional balance approach, first for responses to varying nutrient levels rather than CO2, by growing sunflower seedlings hydroponically over wide ranges of nitrate in solution (3, 10, and 150 micromolar) and of phosphate (0.1, 0.3, 1, and 3 micromolar). We grew plants in solution so that we had exact control and exact measures of the nutrient levels. The levels are very low by agronomic standards; plants grew well despite this because the solution was strongly stirred, allowing roots good access to the nutrient. At the highest levels the plants doubled in size every 3 days, as fast as any plant in any natural or agronomic conditions.
Our plants at different nutrient levels presented notable differences in growth rates (expressed most usefully as relative growth rates per day, RGR), in content of N and of P (as fractions of dry mass, fN and fP), in maximal capacity of roots for uptake of nutrients (Vmax,N and Vmax,P), in an offset in the uptake rate called the Michaelis constant (Km, value inferred only weakly), and in root-to-shoot mass ratio (r). Our challenge was to figure out if how the changes could be adaptive, giving the best growth rate, and how each type of change contributed to that rate. The functional balance argument is given in our published papers (see footnotes at end) and it’s presented at the end, here. In essence, we equated two relative growth rates:
- one limited by nutrient uptake, tied to
- root investment r,
- uptake capacity Vmax,
- solution nutrient content ce and uptake offset Km, and
- “targeted” tissue nutrient content fN)
- and one limited by nutrient use in performing photosynthesis, tied to
- root:shoot ratio r as a “diversion” away from leaves,
- photosynthetic efficacy of the nutrient as photosynthetic rate per mass of nutrient p,
- rfactional investment of shoot mass into leaves αL,
- efficiency of converting photosynthetic sugars into final biomass β, and
- solution nutrient content).
One result is that the tissue N content should vary as the square root of the photosynthetic efficacy of N. With the great effort needed for measuring many other traits we could not measure the photosynthetic rate directly but could infer it from growth rate and leaf area. Our final expression is
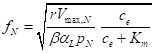
Here ce is the concentration of N as nitrate in the solution. We did least-squares fitting of the natural logarithm of fN (to make the fit linear) and this behavior was supported statistically.
Another expression is for relative growth rate itself:
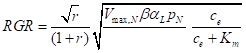
This was also supported statistically.
All the plants were healthy, even at the very low nutrient levels. The net response to lower nutrient levels was to increase the root:shoot ratio (N acquisition is more of a limiting factor than is using it in photosynthesis), increase Vmax for uptake (only at the lowest levels), and decrease nutrient content in the balanced form proposed above.
This formula has its limitations, as the balance of functions in the plant changes with age and ecological setting. At late maturity of an annual plant, for example, nutrient acquisition becomes unimportant. Also, plants may face water limitation more than nutrient limitation at some seasons. Nonetheless, it is a start.
With collaborators Ayalsew Zerihun and Hormoz BassiriRad, I participated in a study of these responses to different CO2 levels. Again, the study plant was the sunflower, Helianthus annuus L. , variety Teddy Bear (I had to note that). The plants were grown from seed and replicates were harvested at two times. We measured growth rates, photosynthetic rates, nitrogen uptake rates, and tissue nitrogen content. The same functional balance model was employed, now with direct measurement of photosynthetic efficacy. The measure of validity of the functional balance model was via plotting predicted bs. observed tissue N content and similarly for relative growth rate. The results are encouraging. There was a shift in offset in N content between harvests, while within a harvest the fit is good. For relative growth rate the fit is even better.
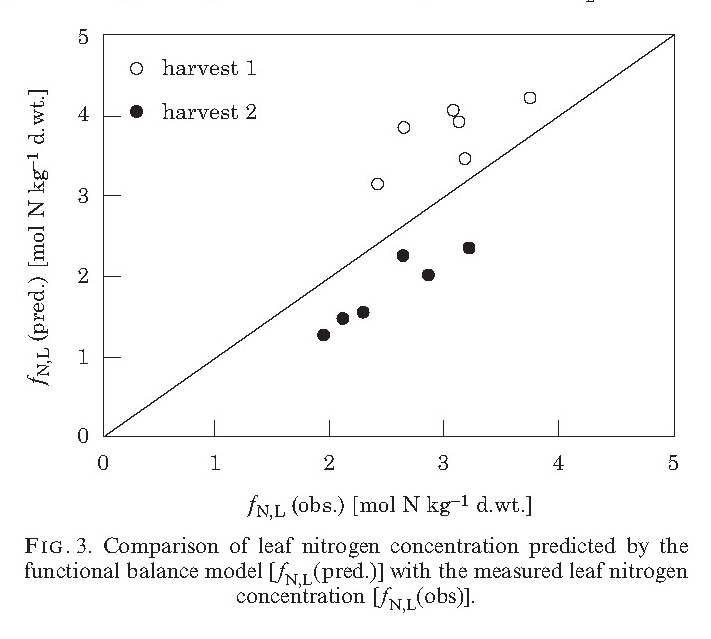

The drop in nutrient content at increasing CO2 levels is of great import ecologically, agronomically, nutritionally, climatologically, and entomologically:
- Ecologically: the competition of plants of different species is very sensitive to differences in growth rates and in nutrient uptake, even to “cornering of the market.” I have made model studies (2003) of changes in species competition as dependent on differences in the kinds of acclimations (individual plants) and adaptations (genetic changes in populations).
- Agronomically and nutritionally: the drop in nitrogen content is a drop in protein content. Durum “hard” wheat for human consumption has to have a high protein content. Crop breeders are having a hard time keeping up with the CO2-induced drop. More generally, the drop in content of various nutrients in a number of human food crops is becoming an issue of concern.
- Climatologically: Which species thrive, which species replace others, makes a difference in several climate processes. For example, families of plants with the C4 mode of photosynthesis have evolved at least 20 times in the past 20 million years. They have a different initial capture of CO2 that gives them a higher efficiency in using nitrogen for photosynthesis and a higher efficiency in using water. Surprisingly they have not yet overrun the world but they are favored in biomes with N and water stress that are variously popping up and disappearing in large regions of the globe. A net change from good old C3 photosynthesis to C4 photosynthesis will change the water flux through land plants and will have add-on effects in precipitation and iwnd patterns.
- Entomologically: Insect populations have decreased an estimated 60% globally in recent decades (Kunin, 2019)! It’s not just insecticides or change of land use. Scientists are examining the role of the decline in plant nutrient content. National Geographic in 2020 had an article that stated we may miss them when they’re gone!
The net effect on Earth is a tuning of habitability for us humans as well as for our fellow inhabitants – plants, insects, mammal, fungi, you name it. The full import of changes in nutrient content and many other traits will play out in ways we have yet to imagine. For other planets that might be habitable, we haven’t a clue about the role of CO2 flutuations… nor even for Earth.
References:
D. VanLeeuwen, Dawn, R. St Hilaire, V. P. Gutschick, Vince P., B. Cook, R. J. Heerema, and V. P. Gutschick. 2014. Leaf photosynthesis in nitrogen-starved ‘Western’ pecan is lower on fruiting shoots than non-fruiting shoots during kernel fill. Journal of the American Society for Horticultural Science 139: 267-274.
V. P. Gutschick. 1993. Nutrient-limited growth rates: Roles of nutrient-use efficiency and of adaptations to increase uptake rate. Journal of Experimental Botany 44: 41-51.
V. P. Gutschick and L. E. Kay. 1995. Nutrient-limited growth rates: quantitative benefits of stress responses and some aspects of regulation. Journal of Experimental Botany 46: 995-1009.
A. Zerihun, V.P.Gutschick, and H. BassiriRad. 2000. Compensatory roles of nitrogen uptake and photosynthetic N-use efficiency in determining plant growth response to elevated CO2:Evaluation using a functional balance model. Annals of Botany 86(4): 723-730.
A. Zerihun, V. P. Gutschick, and H. BassiriRad. 2000. Compensatory roles of nitrogen uptake and photosynthetic N-use efficiency in determining plant growth response to elevated CO2: Evaluation using a functional balance model. Annals of Botany 86: 723-730.
V. Gutschick. 2006. Plant acclimation to elevated CO2—From simple regularities to biogeographic chaos. Ecological Modelling 200: 433-451.
W. E. Kunin, 2019. Robust evidence of declines in insect abundance and biodiversity. Nature 574, 641-642.